DNA-reparasjons mekanismer designet med spesiell forsiktighet for å gi integritet for DNA, og essensielt for levende organismer av alle domener
Av Otangelo Grasso
Oversatt herfra.
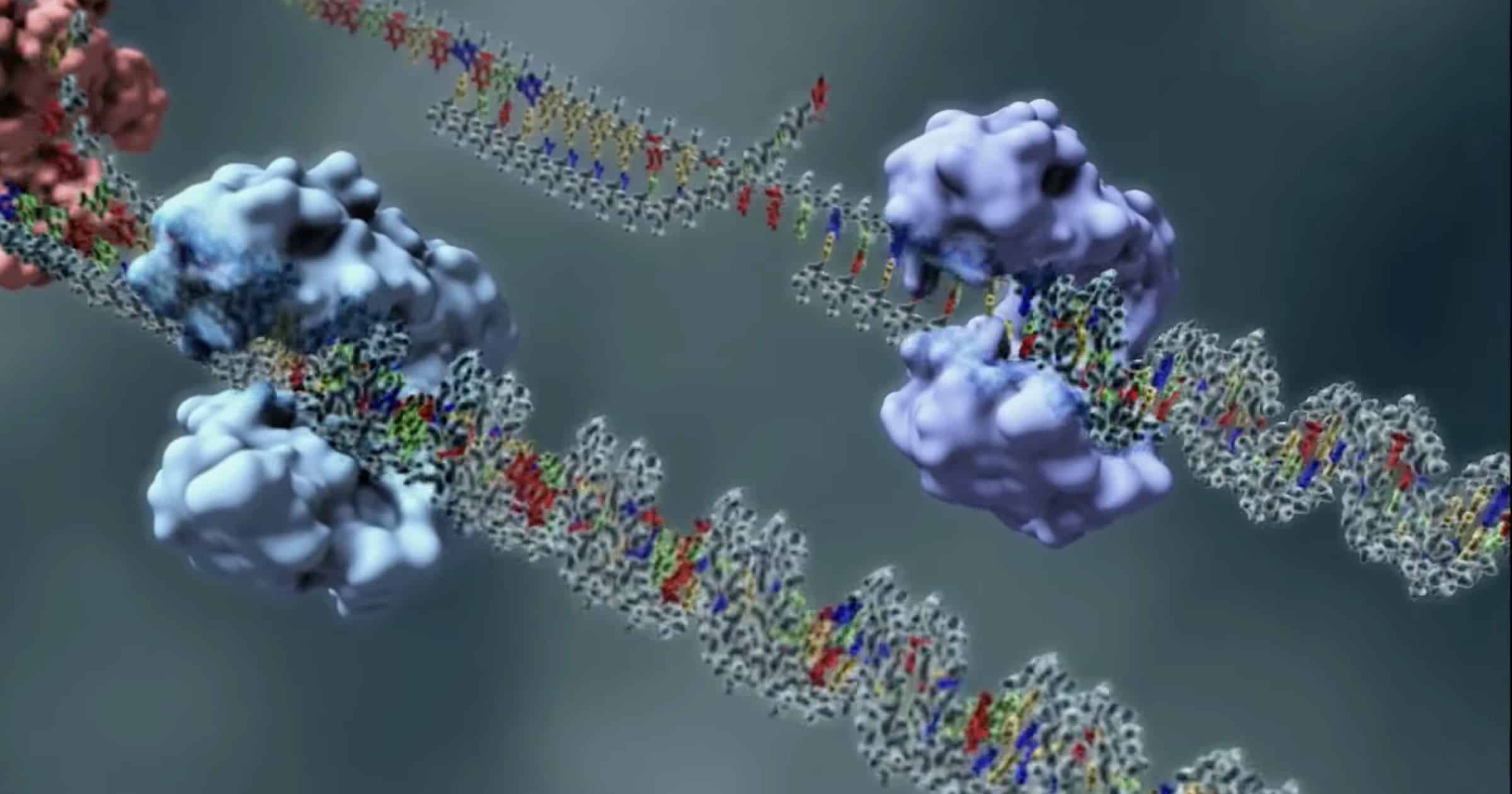 Å opprettholde den genetiske stabiliteten som en organisme trenger for å overleve, krever ikke bare en ekstremt nøyaktig mekanisme for replikering av DNA, men også mekanismer for å reparere de mange tilfeldige lesjonene som oppstår kontinuerlig i DNA.
Å opprettholde den genetiske stabiliteten som en organisme trenger for å overleve, krever ikke bare en ekstremt nøyaktig mekanisme for replikering av DNA, men også mekanismer for å reparere de mange tilfeldige lesjonene som oppstår kontinuerlig i DNA.
DNA-skade er en endring i den kjemiske strukturen til DNA, for eksempel et brudd i en DNA-streng, en base som mangler fra ryggraden til DNA, eller en kjemisk endret base. Naturlig forekommende DNA-skader oppstår mer enn 60 000 ganger per dag per pattedyrcelle. DNA-skader ser ut til å være et grunnleggende problem for livet. DNA-skader er en hovedårsak til kreft. DNA-skader gir opphav til mutasjoner og epimutasjoner. Mutasjonene, hvis de ikke korrigeres, ville bli forplantet gjennom påfølgende cellegenerasjoner. En så høy grad av tilfeldige endringer i DNA-sekvensen ville ha katastrofale konsekvenser for en organisme.
Bilde 1. Eks. på reparasjonsverktøy (Topoisomerase)
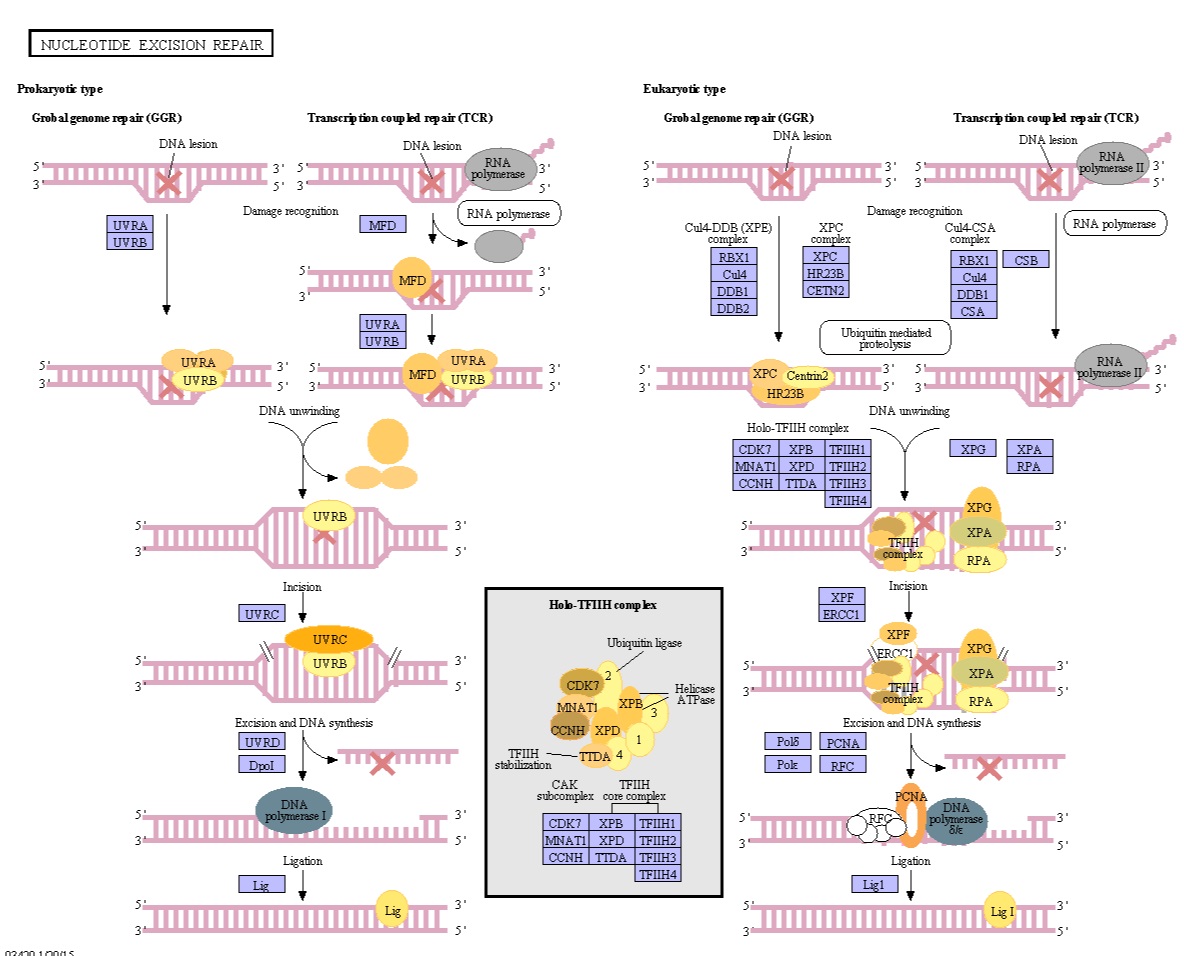
Ulike veier for DNA-reparasjon eksisterer, nukleotideksisjonsreparasjon (NER), baseeksisjons-reparasjon (BER), DNA mismatch reparasjon (MMR), reparasjon gjennom alkyltransferase-lignende proteiner (ATLs) blant andre.
Det er tydelig at reparasjonsmekanismen er avgjørende for at cellen skal overleve. Det kunne ikke ha utviklet seg etter at livet oppsto, men må ha blitt til før. Mekanismen er svært kompleks og utdypet, som en konsekvens er designslutningen berettiget og ser ut til å være den beste måten å forklare dens eksistens på.
Base excision repair (BER) involverer en kategori av enzymer kjent som DNA-N-glykosylaser.
Ett eksempel på DNAs automatiske feilrettingsverktøy er nok til å forvirre fantasien. Det finnes dusinvis av reparasjonsmekanismer for å beskytte vår genetiske kode mot skade; en av dem ble fremstilt i Nature i termer som burde vekke ærefrykt.
Fra Natures artikkel:
Strukturen til et reparasjonsenzym som undersøker uskadet DNA, belyser gjenkjennelse av skadet DNA (11)
Hvordan DNA-reparasjonsproteiner skiller mellom de sjeldne skadestedene og det store området med normalt DNA er dårlig forstått. Å gjenkjenne den mutagene lesjonen 8-oksoguanin (oxoG) representerer en spesielt formidabel utfordring, fordi denne oksiderte nukleobasen skiller seg med bare to atomer fra sin normale motpart, guanin (G). Røntgenstrukturen til det fangede komplekset har en mål-G-nukleobase ekstrudert fra DNA-helixen, men nektet innsetting i lesjonsgjenkjenningslommen til enzymet. Beregninger av fri energiforskjell viser at både attraktive og frastøtende interaksjoner har en viktig rolle i den foretrukne bindingen av oxoG sammenlignet med G til det aktive stedet. Strukturen avslører en bemerkelsesverdig effektiv portholdingsstrategi for lesjonsdiskriminering og foreslår en mekanisme for oxoG-innsetting i det aktive hOGG1-stedet.
Bilde 2. DNA-reparasjoner behøves ofte (Illustrasjon fra O. Grasso)
 Av de fire basene i DNA (C, G, A og T) er cytosin eller C alltid ment å parre med guanin, G, og adenin, A, er alltid ment å parre med tymin, T. Enzymet studert av Banerjee et al. i naturen er en av en rekke molekylære maskiner kalt BER-glykosylaser; denne kalles humant oxoG-glykosylase-reparasjonsenzym (hOGG1), og den er spesialisert for å finne en bestemt type feil: en oksidert G-base (guanin). Oksidasjonsskader kan være forårsaket av eksponering for ioniserende stråling (som solbrenthet) eller frie radikaler som streifer rundt i cellekjernen. Den normale G blir oxoG, noe som gjør den veldig lite, ute av sin form. Det kan være én av en million av disse på en DNA-streng. Selv om det virker som en mindre skrivefeil, kan det faktisk føre til at oversettelsesmaskineriet setter inn feil aminosyre i et protein, med katastrofale resultater, for eksempel tykktarmskreft. (12)
Av de fire basene i DNA (C, G, A og T) er cytosin eller C alltid ment å parre med guanin, G, og adenin, A, er alltid ment å parre med tymin, T. Enzymet studert av Banerjee et al. i naturen er en av en rekke molekylære maskiner kalt BER-glykosylaser; denne kalles humant oxoG-glykosylase-reparasjonsenzym (hOGG1), og den er spesialisert for å finne en bestemt type feil: en oksidert G-base (guanin). Oksidasjonsskader kan være forårsaket av eksponering for ioniserende stråling (som solbrenthet) eller frie radikaler som streifer rundt i cellekjernen. Den normale G blir oxoG, noe som gjør den veldig lite, ute av sin form. Det kan være én av en million av disse på en DNA-streng. Selv om det virker som en mindre skrivefeil, kan det faktisk føre til at oversettelsesmaskineriet setter inn feil aminosyre i et protein, med katastrofale resultater, for eksempel tykktarmskreft. (12)
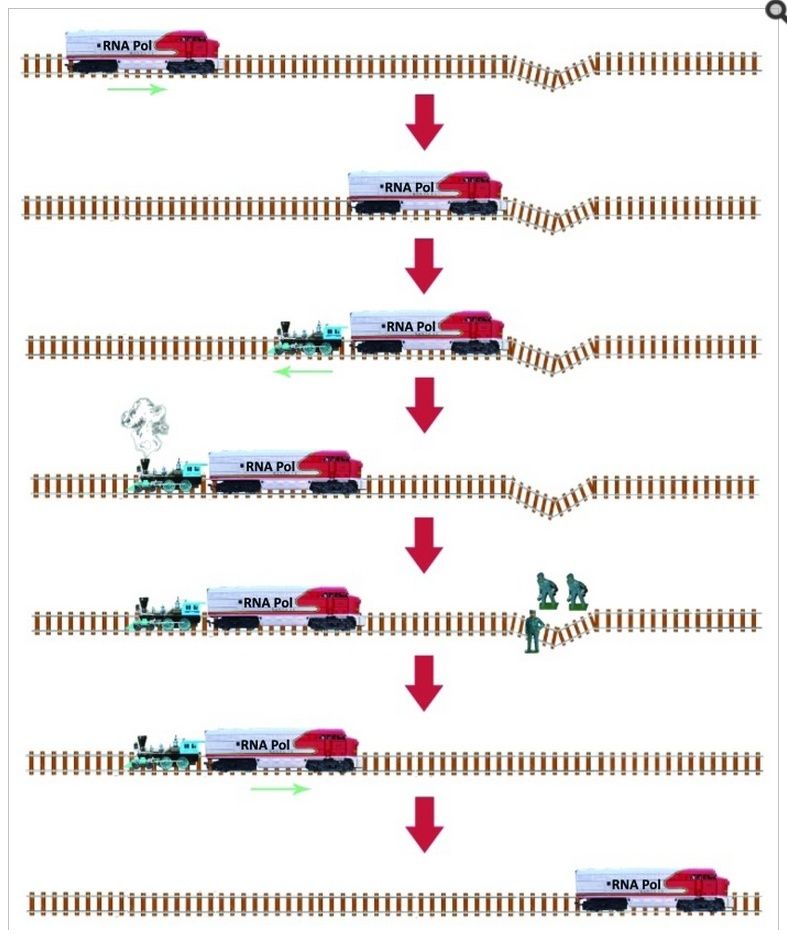
Maskinen låses fast på DNA-dobbelthelixen og jobber seg nedover tråden, og kjenner hver base på veien. Når den fortsetter, knekker den DNA-tråden til en skarp vinkel. Den er bygget for å ignorere T- og A-basene, men når den føles som en C, vet den at det skal være en G festet. Maskinen har presisjonskontaktpunkter for C og G. Når C-en kobles inn, vippes basen som er sammenkoblet med den, opp ut av helixen og inn i en spalte inne i enzymet som er fint laget for å pare seg med en ren, ren G. Hvis alt er vel, snur den G-en tilbake i DNA-helixen, og går videre. Hvis basen er en oxoG, blir imidlertid basen snudd inn i et annet spor lenger inne, hvor kraftige krefter drar den feilaktige basen ut av strengen slik at andre maskiner kan sette inn den riktige.
Nå er alt dette fantastiske så langt, men som med mange ting i levende celler, ligger det sanne underet i detaljene. De termodynamiske energiforskjellene mellom G og oxoG er ekstremt små – oxoG inneholder bare ett ekstra oksygenatom – og likevel er denne maskinen i stand til å skille mellom dem med høy nøyaktighet.
 Forfatteren, David, sier i Nature-artikkelen: Strukturell biologi: DNA-søk og redning
Forfatteren, David, sier i Nature-artikkelen: Strukturell biologi: DNA-søk og redning
DNA-reparasjonsenzymer forbløffer oss med deres evne til å søke gjennom store DNA-områder for å finne subtile anomalier i strukturen. Det menneskelige reparasjonsenzymet 8-oksoguaninglykosylase (hOGG1) er spesielt imponerende i denne forbindelse fordi den fjerner effektivt 8-oksoguanin (oxoG), en skadet guanin (G)-base som inneholder et ekstra oksygenatom, og ignorerer uskadede baser.
Naturlig seleksjon kan ikke handle uten nøyaktig replikering, men proteinmaskineriet for nøyaktighetsnivået som kreves, er i seg selv bygget av den genetiske koden den er designet for å beskytte. Det er en catch22-situasjon. Det ville vært utfordrende nok å forklare nøyaktig transkripsjon og oversettelse alene med naturlige midler, men som følge av UV-stråling ville den raskt blitt ødelagt gjennom akkumulering av feil. Så nøyaktig replikering og korrekturlesing er nødvendig for livets opprinnelse. Hvordan i all verden kunne korrekturlesende enzymer dukke opp, spesielt med denne grad av troskap, når de er avhengige av selve informasjonen de er laget for å beskytte? Tenk på det.... Dette er enda et prima facie eksempel på kylling- og eggsituasjon. Hva er den alternative forklaringen på design? Korrekturlesing av DNA ved en tilfeldighet? Og en kompleks serie med oversettelsesmaskineri uten designer?
Bilde 3. Illustrasjon av DNA-reparasjon (fra O. Grasso)
Jeg liker å lære om underverket med disse utrolige mekanismene. Hvis apostelen Paulus kunne forstå at skapelsen krever en Skaper som han skrev i Romerbrevet kapittel 1 18, hvor mye mer vi i dag med alle åpenbaringene om cellebiologi og molekylære maskiner?
Oversettelse ved Asbjørn E. Lund